
关于太湖摇蚊幼虫对底质环境及内源释放影响的研究
作者:曹健国
据报道,摇蚊幼虫和颤蚓类的生命活动能促进沉积物中氮以铵的形式向水体大量释放,但对于磷的释放没有研究定论,且作用机制尚不明确,特别是生物扰动对泥水界面的氧化还原环境、沉积物复氧的影响机制是什么,尚缺乏系统的研究和验证。基于此,本研究选取太湖富营养化湖区梅梁湾,采集柱状样进行原位静态培养模拟实验,探讨和比较主要优势种类羽摇蚊和长足摇蚊的生物扰动对底质环境的再造作用,以及对内源释放的作用机制。以期深入掌握富营养化湖泊的内源发生机制及其生物扰动效应,为控制水体内源污染释放和富营养化治理提供科学依据。
1 湖泊概况和研究方法
1.1湖泊概况
太湖梅梁湾(Meiliang Bay)位于太湖北部,是一个半封闭的湖区,面积100 km2,平均水深2.2 m,透明度0.2 m左右,该湖区接纳了来自常州和武进等地的城市污水和工业废水,水质N~V类,属重污染藻型湖区,夏季易暴发水华,局部黑臭。
1.2生物扰动效应的模拟试验
1.2.1泥一水系统的建立
本实验于梅梁湾湖心用装有φ90 mmx500 mm有机玻璃管的柱状采泥器采集18根原位柱状样,泥样厚度26 cm,上部用原样点水样注满后两端用橡皮塞塞紧,垂直放置,小心运回实验室以进行原位静态培养试验。试验开始前,将柱状样中上层水体用虹吸法抽去,再用虹吸法沿壁小心滴注经GF/C膜过滤的上覆水样(除去藻类)至液面高度20 cm停止,整个过程避免扰动界面,建立柱状泥水系统后静置,待泥水系统平衡24 h后开始试验。
1.2.2试验条件
将柱状泥水系统置于地下恒温室((22±1)℃),避
光静态培养。泥水系统平衡24 h后进行试验。
1.2.3试验处理
本研究选取梅梁湾底栖动物的优势种类羽摇蚊Chironomus plumosus和长足摇蚊Tanypus chinensis的幼虫作为试验动物,试验前挑选个体较大、健壮、活动能力较强的虫体(羽摇蚊虫体长度15~16 mm、长足摇蚊虫体长度8~10 mm),进行室内恒温培养驯化24 h,尔后添加至上述柱状泥水系统中进行生物扰动模拟试验。试验处理为添加羽摇蚊、添加长足摇蚊和对照不添加任何底栖动物,添加密度均为1 200条/m2,每个处理设3个平行柱状样。
1.2.4试验方法
试验系统分为2组,一组柱状泥水系统(9根)用于进行营养释放试验,另一组柱状泥水系统(9根)用于进行沉积物耗氧试验,2组试验同时进行,处理和平行相同。
营养释放试验具体步骤如下:添加虫子后,分别于O、12、24、48、72、120 h取上覆水样50 mL测定溶解无机态氮( NH4+-N,N02- -N,N03- -N)、溶解反应性磷(SRP),每次取样后用原样点初始过滤水样补充同体积水样。同时,用YSI 550A溶氧仪测定系统上覆水中的溶氧量。120 h后,抽去上覆水,底泥以2 cm/层分层切割后用于测定沉积物含水率、间隙水中溶解态无机氮、磷含量以及微生物FDA活性和碱性磷酸酶APA活性。
溶解态无机氮、磷的测定按《水和废水监测分析方法》进行;沉积物含水率为沉积物烘干前后重量差值与原有湿沉积物重量的比值;微生物活性采用FDA(荧光素双醋酸酯)法,测定步骤如下:称取1.00 g左右湿沉积物样,加入10 mL灭菌后pH=7.0的0.01mo1/L的磷酸缓冲溶液,于30℃、200 r/min摇床中分散15 min,之后加入2 g/L的FDA液0.5 mL,于30℃摇床中显色3h,离心、过滤,于490 nm波长处比色,测定吸光度,换算成单位质量干泥的吸光度值,以此来表示沉积物中微生物FDA活性;碱性磷酸酶的测定方法:取一定量的湿沉积物,加入pH=8.6的0.5 mol/L的Tris-HCl缓冲液,以1:2的比例混匀制成匀浆,取2 mL匀浆,加入一定浓度底物(p-NPP)0.5 mL,放入恒温37℃摇床中培养th后放入冰柜中冷却,然后分别加入0.5 mol/L的CaCl2、NaOH 0.5 mL和2 mL终止反应,离心,410 nm比色,换算成单位质量干泥的活性强度。
沉积物耗氧(SOD)试验方法如下:试验前用虹吸法取上覆水100 mL测定溶氧初始值,尔后加水至顶端塞紧橡皮塞至有水溢出即密封,密封静置4h后取样分析溶氧量,通过计算密封前后上覆水溶氧量的差值获得沉积物耗氧量(SOD),溶氧量的测定采用碘量法进行。
1.3计算与分析
营养盐的释放速率按式(1)计算:

式(1)中,R为释放速率(mg/(m2·d));V为柱样中上覆水体积(L);Gn、Co、Cj-1分别为第n次、初始和第j-l次采样时某物质含量(mg/L);Ca为添加原水中的某物质含量(mg/L);Vj-l为第j-l次采样体积(L);A为柱样中水一沉积物接触界面(m2);f为释放时间(d)。由于不考虑NH3的水气界面交换,所计算的NH4+-N为表观释放速率。
参数的显著性检验在Statistic 6.0上进行。
2研究结果
2.1 对底质环境的生物扰动效应
2 .1.1对上覆水溶氧的影响
如图1所示,各处理组上覆水的溶氧量均随时间呈降低趋势,培养48 h后羽摇蚊和长足摇蚊处理组的上覆水溶氧量均显著低于对照组(p<0.05),培养时间120 h达最低值,分别为(1.5±0.4) mg/L和( 2.3±0.4) mg/L,羽摇蚊和长足摇蚊处理间差异不显著(p>0.05)。

2 .1.2对沉积物理化环境的影响
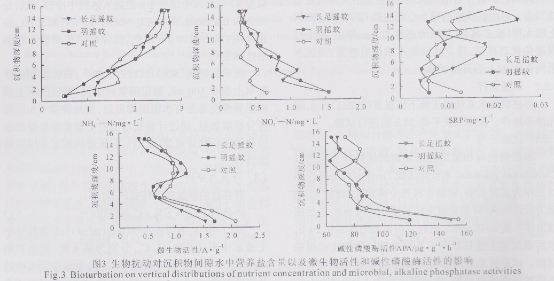
如图2所示,从沉积物含水率的垂直分来看,各处理组沉积物含水率均随深度递增而呈降低趋势,处理间差异不明显。从沉积物耗氧量来看,添加幼虫的沉积物耗氧量均显著高于对照(p<0.05),幼虫间差异不显著。培养试验结束后,从沉积物间隙水中溶解态无机氮、磷的垂直分布来看(图3),各处理组氨氮( NH4+-N)浓度均随深度递增而呈上升趋势,处理间差异不显著。在底泥0~8 cm深处,添加幼虫处理组的底泥间隙水中硝态氮(NO3--N)含量显著高于对照(p<0.05)。长足摇蚊处理组的溶解反应性磷(SRP)含量在底泥5~10 cm深处急剧上升至0.02 mg/L,12~14 cm深处达最大值0.025 mg/L,显著高于对照和羽摇蚊处理组( p<0.05),其他处理间差异不显著(p>0.05)。从底泥微生物活性和碱性磷酸酶活性的垂直分布来看(图3),各处理间的微生物活性差异不显著;长足摇蚊处理组的磷酸酶活性在沉积物8~10 cm深处显著高于对照组,而在10~12 cm深处,羽摇蚊处理组的磷酸酶活性显著低于对照组(p<0.05),其他深度的处理间差异不显著(p>0.05)。
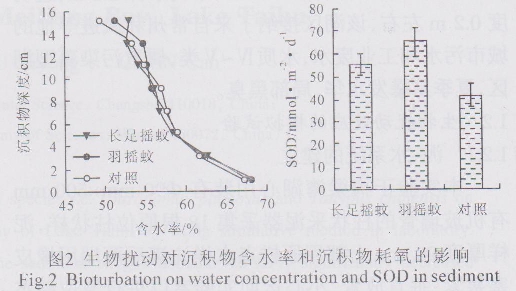
2.2对内源释放的影响
如图4所示,羽摇蚊和长足摇蚊均促进了沉积物氨氮向上覆水的释放,释放速率分别为( 20.5±9.3)mg/(m2.d)和(18.3±2.4) mg/( m2.d),显著高于对照(p<0.05);各处理组沉积物硝态氮均随时间呈负释放趋势,羽摇蚊处理组硝态氮的释放速率为( 48.6±0.7)mg/(m2·d),显著高于对照(37.3±3.0)mg/( m2·d),120 h羽摇蚊处理组的上覆水中硝态氮含量达到最低值,显著低于对照(p<0.05),长足摇蚊处理组硝态氮的释放速率为(43.6±2.1)mg/( m2.d),与对照差异不显著(p>0.05);各处理组溶解反应性磷随时间呈释放趋势,羽摇蚊处理组溶解反应性磷的释放速率为( 0.96±0.05) mg/( m2·d),长足摇蚊处理组为( 0.88+0.08) mg/( m2·d),对照组为( 1.01±0.10)mg/( m2·d),处理间差异不显著(p>0.05)。

3讨论
底栖动物在沉积物中的呼吸、摄食、爬行和筑巢等行为,及其对营养盐的吸收、同化和排泄活动能加快泥水界面的物质交换,对泥水界面氮、磷元素的生物地球化学循环产生重大影响。本研究通过开展扰动效应的室内模拟,深入比较了2种摇蚊幼虫(羽摇蚊和长足摇蚊)对泥水界面的理化环境以及内源释放的扰动效应。羽摇蚊生活于沉积物中5~20 cm深处,以有机碎屑为食,具筑巢穴的生态习性。本研究观察发现沉积物表层至底层均可见有明显的爬行痕迹和穴管通道。与羽摇蚊相比,长足摇蚊个体小,活动能力弱,据观察,幼虫主要在沉积物表层2 cm左右深处活动,无筑巢现象。据报道,底栖动物,如摇蚊幼虫,水蚯蚓的物理扰动和引灌作用能促进沉积物的复氧和耗氧作用,同时降低泥水界面的溶氧量,本研究亦发现,羽摇蚊和长足摇蚊均促进沉积物的耗氧作用(图2),降低了上覆水的溶氧量,改变了泥一水界面的氧化还原条件,从而对界面氮、磷的生物地球化学循环产生了重大影响。矿化和形态转换均为氮循环的主要过程,其中影响氮形态转换的2个重要微生物过程包括硝化作用和反硝化作用。在通气良好的条件下,铵(或氨)被硝化细菌作用而氧化成硝酸的过程叫做硝化作用。硝酸根在嫌气(氧气不足)条件下被反硝化细菌作用而还原成一氧化二氮或氮气的反应叫做反硝化作用。有研究表明,底栖动物生活的巢穴和爬行的管道等构造具较强的硝化作用潜力,原因在于这些构造中有充足的NH4+和氧气来源。本研究发现羽摇蚊和长足摇蚊的生物引灌作用均增加了沉积物的通透性,特别是氧气的通透作用,促进了沉积物发生硝化作用,从而导致沉积物硝态氮的溶出(以间隙水中硝态氮含量显著高于对照为证,图3)。同时,羽摇蚊的生物扰动和呼吸代谢大大降低了上覆水的溶氧量,从而促进上覆水发生反硝化,是羽摇蚊处理组上覆水中硝态氮含量显著低于对照的重要因素(图4)。2种幼虫的扰动均显著促进了氨氮的释放(图4),究其原因在于:幼虫的生物引灌作用加速了沉积物的耗氧和矿化作用(以沉积物耗氧量显著增长为证,图2),以及代谢产物向上覆水的扩散。此外,幼虫自身的生理代谢排铵作用亦不可忽略。
而生物扰动作用对磷释放的影响机制较为复杂,与扰动生物的种类、习性和底泥的性质有关,已有研究结果存在争议,有研究认为羽摇蚊的生物扰动作用促进了磷酸盐的释放,认为虫子的生理代谢作用是磷酸盐释放的重要因素;而Lewandowski、Geraldine等的研究结果表明羽摇蚊的生物扰动抑制了磷的释放,认为生物引灌增加了沉积物的氧化性,从而促进沉积物中铁磷的吸附,是抑制磷释放的重要因素;Filip等的研究结果表明,羽摇蚊对磷的释放作用不显著,本研究得到了同样的结果。而关于长足摇蚊的生物扰动效应研究甚少,未见相关报道。本研究发现长足摇蚊的生物扰动能促进沉积物碱性磷酸酶的活性,从而促进沉积物磷酸盐向间隙水的溶出(图3)。由此看来,生物扰动对磷释放的效应取决于以上因素的综合结果。
本研究通过室内模拟的方法系统地探讨了富营养化湖泊中常见优势种类羽摇蚊和长足摇蚊的生物扰动对底质环境以及泥一水界面氮、磷循环的影响机制。研究结果表明:2种摇蚊幼虫通过各自独特的物理扰动和生理代谢方式,改变了泥一水界面的理化环境,集中表现在通过改变泥水界面的氧化还原条件,以及沉积物耗氧矿化作用来促进界面氮、磷元素的生物化学过程,加速了内源的释放,主要是氨氮的释放。同时对沉积物一间隙水营养盐的溶出和吸附平衡造成重大影响,集中表现在增强了碱性磷酸酶活性,从而促进磷酸盐的溶出,以及生物引灌增强沉积物的通透和复氧,从而促进沉积物发生硝化作用,导致硝态氮向间隙水的溶出。总而言之,2种摇蚊幼虫对富营养化湖泊水一土界面氮、磷元素的循环,特别是内源的释放产生了重大影响。本研究所得结果可为富营养化湖泊的治理,特别是内源负荷的控制提供科学依据。
4摘要:
该研究通过开展室内原位静态模拟试验,探讨了太湖梅梁湾常见优势种类羽摇蚊和长足摇蚊对底质环境以及内源释放的生物扰动效应,研究结果表明:2种幼虫均促进了沉积物的耗氧作用、显著降低了上覆水的溶氧量、促进沉积物中硝态氮向间隙水的溶出、促进沉积物氨氮向上覆水的释放;长足摇蚊促进了沉积物碱性磷酸酶活性和溶解反应性磷的溶出。2种摇蚊幼虫的物理扰动和生理代谢活动改变了泥水界面的氧化还原环境,促进沉积物的耗氧矿化作用,从而加速内源释放,对沉积物一间隙水营养盐的溶出和吸附平衡产生显著影响。
上一篇:关于食品接触材料中荧光增白剂VBL禾口APC迁移规律的探索
下一篇:返回列表